January 2023
Research Trip to Nairobi, Kenya! < see the blog post all about it here in which I travelled to the NMK in Kenya to collect 3D scans of fossil specimens!

December 2022
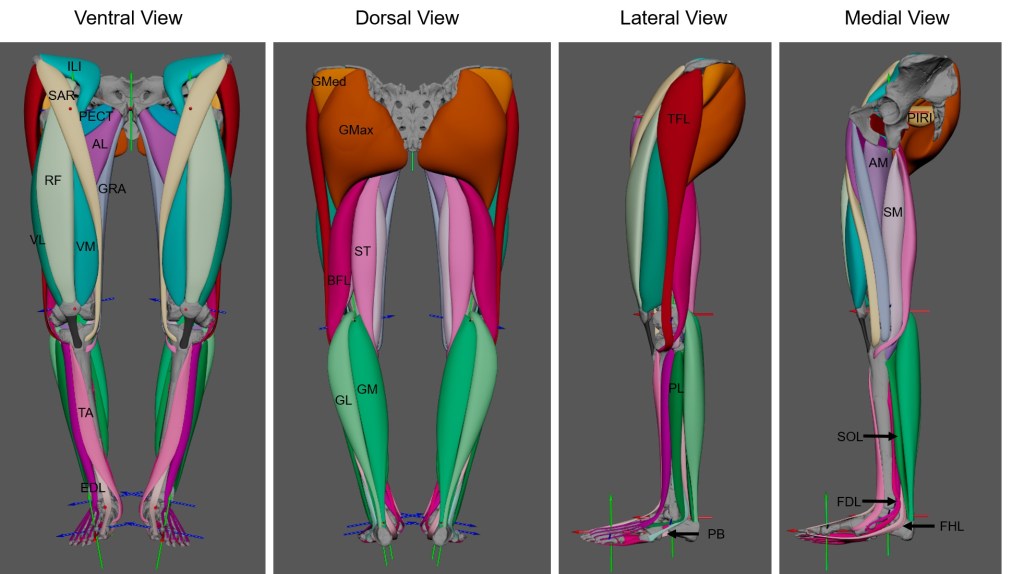
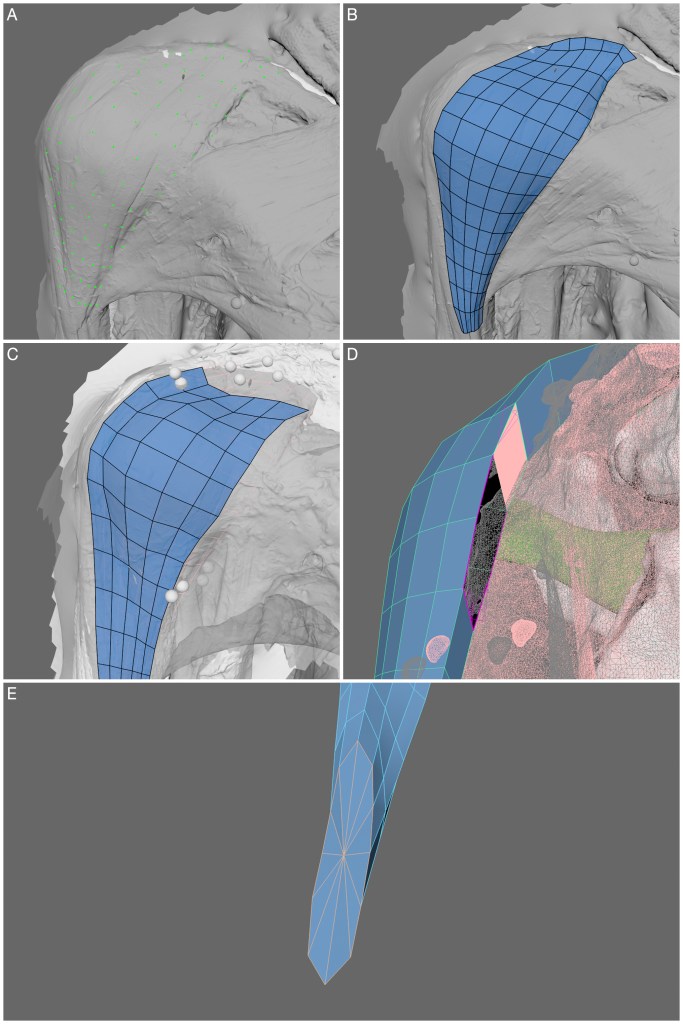
Preprint now available on BioRxiv which details a 3D polygonal muscle modelling approach in the Australopithecus afarensis specimen AL 288-1! See the blog post here about the paper, which also includes videos of the muscles from superficial to deep layers! I had a lot of fun creating these muscles.
So, why do we need to use such an approach? Musculature simply does not survive in the fossil record. Rather, we are left looking at just the bones. But muscles animate movement, allowing an organism to walk, run, jump and even dance. So if we want to understand how an extinct specimen might have moved, then we need to understand its musculature. Typically, palaeontologists/palaeoanthropologists use a method called the Extant Phylogenetic Bracket to accomplish this task which involves identifying the closest living analogy to the extinct specimen to estimate where muscles attached and how they lay in the body. In the past, this produced simple straight lines of action from a point of origin to the insertion location, which fails to capture how the muscle might have wrapped around bones. In the last 20 years, wrapping surfaces and via points have been developed in biomechanical modelling software to target this problem. The 3D polygonal modelling approach expands upon this problem even further by considering how much space a muscle might have occupied and its path within the body (i.e., this muscle lies on top of this muscle and extends in this direction, so forth). To accomplish this, scaled MRI cross sectional scans were used to define muscle boundaries, allowing the creation of 3D muscles which occupy a realistic space within the body.
This is a necessary precursor to future dynamic research. This 3D modelling approach shows great promise for recreating the musculature of other hominins… stay tuned!
November 2022
I will be heading to the Computational Biomechanics Lab at the TU Delft, Netherlands for a few weeks to chat research, dynamics and simulations….
Keep posted!
August 2022
Reconstructing articular cartilage in the Australopithecus afarensis hip joint and the need for modelling six degrees of freedom
The first paper of my Leverhulme Trust funded project has just been published in Integrative Organismal Biology – read it here! So what is our paper about?
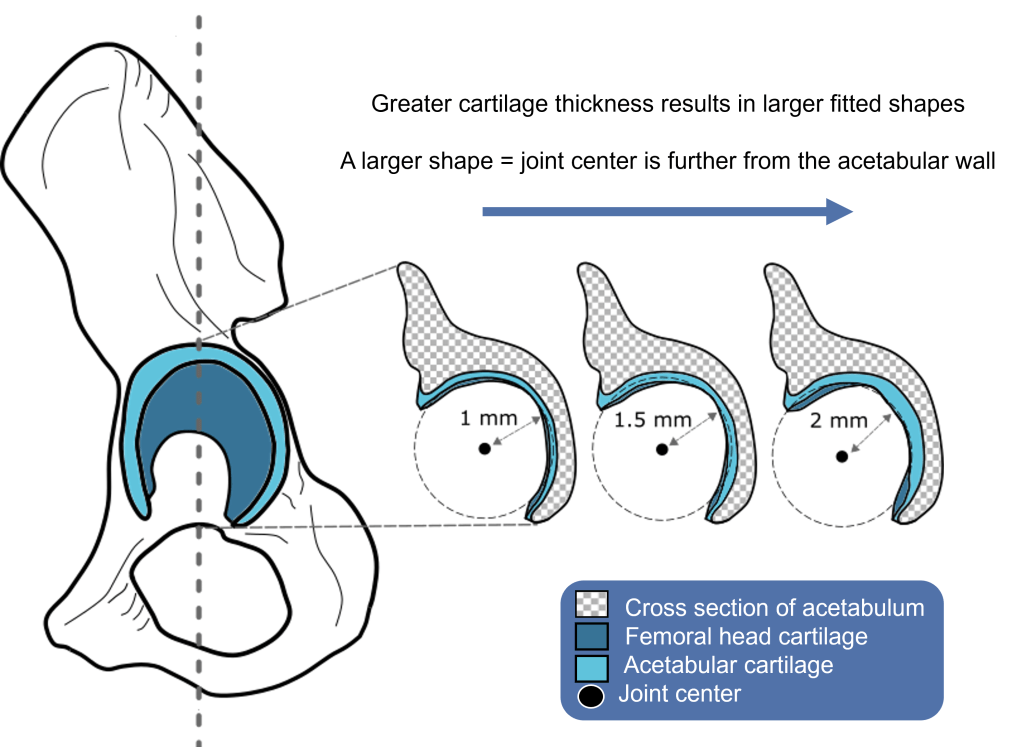
It is unusual for soft tissues to preserve in the fossil record and, therefore, cartilage is very poorly preserved. In the case of the famous Australopithecus afarensis AL 288-1 skeleton (nicknamed ‘Lucy’ after the excavators found her whilst listening to Lucy in the sky with diamonds by The Beatles – read more about the discovery here), around 40% of her skeleton has amazingly been found – this is one of the most complete hominin skeletons discovered to date. But only the hard tissues have preserved – that is, the bones. In each of the joints of a body, articular cartilage is preserved which helps with the dissipation of stress during joint movement. Cartilage can be thick or thinly distributed through the joint’s articular surface (or a mixture of both) depending on the joint and its functionality. So if no cartilage has preserved belonging to Lucy, how can we accurately rearticulate her skeleton? And if we under- or over-predict the joint spacing, what repercussions could this have for claims regarding movement?
The postcranial skeleton of AL 288-1 exhibits clear adaptations for bipedality (that is, it shares many morphological features with humans), although there is some debate as to the efficiency and frequency of such upright movement. Some researchers argue that AL 288-1 walked with an erect limb like modern humans do, whilst others advocate for a “bent-hip bent-knee” gait (similar to how a chimpanzee can walk bipedally), although in recent years the general consensus favours erect bipedalism.
ROM mapping methods have been developed in recent years for a select range of extinct and extant species to ascertain how two body segments articulate and move relative to one another. ROM mapping relies upon movement of a body segment around a joint center and can encompass rotational and/or translational movement. The method identifies which poses are viable and which are non-viable based on bone morphology, thus providing information regarding limb posture. To date, this method has not yet been applied to humans nor the hominin fossil record, but offers potential to address questions regarding the bipedal gait of early hominins, such as the probable biped Australopithecus afarensis.
And this is where our ROM mapping (Range of Motion) approach comes into play! We employed ROM mapping methods to estimate the joint spacing of AL 288-1’s hip joint in comparison to a modern human and chimpanzee. Nine simulations assessed different joint spacing and tested the range of joint congruency (i.e., ranging from a closely packed socket to loosely packed). We further evaluated the sphericity of the femoral head and whether three rotational degrees of freedom (DOFs) sufficiently captures the full ROM or if translational DOFs must be included.
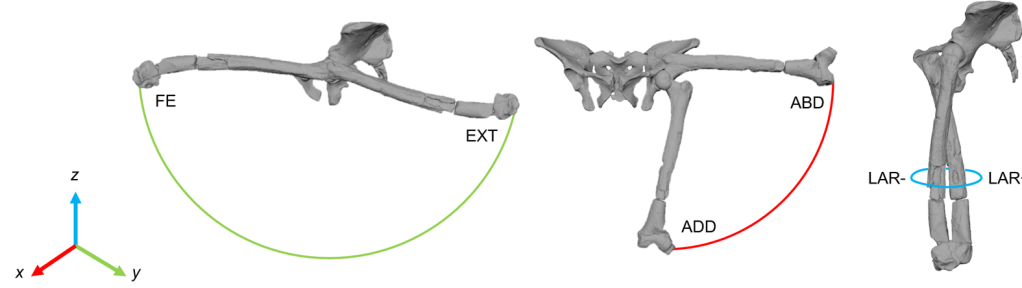
But what do we mean by rotational versus translational? Well, every joint can move in different ways. If you stand up and lift your leg upwards, that is flexion. Moving it backwards is extension. You can move your leg away from the midline of your body (abduction), and can cross you thigh in front of your other leg (adduction). You can also rotate your leg inwards and outwards. That is what we mean by rotational movement. But movement is dynamic and fluid, and doesn’t occur from a central pivot point. Rather, your joint centre can move within a constrained space, thus permitting a greater range of motion. That is what we mean by translational. Some types of ROM studies only test motion using rotational DOFs (DOFs = flexion/extension, or abduction/adduction, or external/internal rotation, or even translation along a single axis etc), but more recent research has established the requirement for translational DOFs to be included, otherwise a joint’s true mobility is underestimated (see here).
Example of rotational movements in the AL 288-1 hip
How did we do this? A full overview of the methods can be read in the paper, but in brief we used the pelvis and femur of a (1) human, (2) chimpanzee and (3) Lucy. We used a shape fitting procedure to establish DOFs, axes and joint centres:
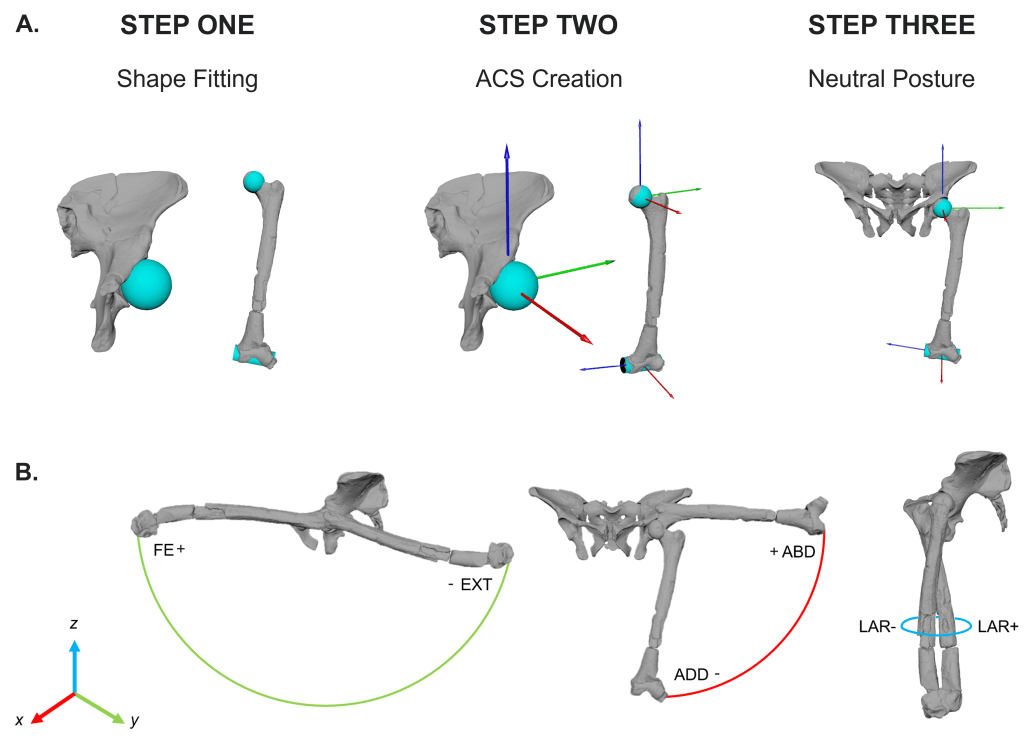

Next, we measured cartilage thickness in chimpanzee and human MRI/CT scans and established a minimum, maximum and average thicknesses of the joint. We applied the human and chimpanzee form (scaled by femoral length) to the Lucy hip joint (simulations 1-6) to establish how rotational-only movement is influenced by joint spacing. To accomplish this, we performed a static Y-axis translation, thereby moving the femoral head further away from the acetabular wall. Interestingly – the simulation which used just the fitted shapes (a proxy for predicting spacing in extinct species) produced zero viability – this was our first indication that translational DOFS should be incorporated. Other rotational-only simulations predicted that AL 288-1 was unable to extend the hip into positions necessitated by bipedal walking!! That would mean that AL 288-1 would have to have walked with a bent hip bent knee when walking upright, like a chimpanzee would do. But this is problematic and extremely unlikely because it contradicts 40+ years of biomechanical research into this species’ movement!
So, what does this mean? Well, it means that there is an issue with the setup. Three DOFs are insufficient to capture true mobility of the hominin hip.
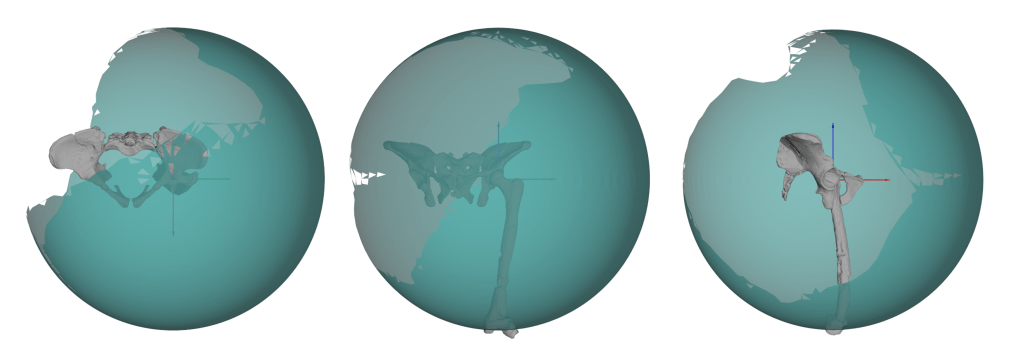
Next, we included translational DOFs. The joint centre was able to move around a sphere which was created inside the acetabulum (all code + instructions to create and implement a sphere-approach are freely available to download as SI). The radius of the sphere was set to the difference of the fitted shapes of the acetabulum and femoral head = 2.448 mm. Interestingly, this value was also highlighted in the results of the three DOF simulations as being the most viable (that is, most functionality was present at this threshold). In this translational setup, AL 288-1 was capable of moving the femur into a range of possible osteological limb poses in the same way as both a chimpanzee and a human – which we concluded as being much more realistic.
To sum up (TLDR): We tested the articulation and possible osteological ROM of the AL 288-1 hip joint by modelling a static single axis translation to investigate increasing joint spacing, which was considered a proxy for measuring maximum cartilage thickness. We expanded upon this by including all six DOFs, thereby reflecting true joint movement. Whilst the resultant ROM maps were quite similar, there was a greater spectrum of viability in the six DOF simulation than the other simulations, in which the femur was capable of osteologically moving into a greater range of poses. With this spectrum of poses, AL 288-1 was capable of a repertoire of movements, such as erect bipedalism across a range of substrates at various speeds and vertical climbing. Overall, six DOFs are a requirement for modelling mobility in fossil hominins, otherwise the resultant functionality of a given joint may be wrong.
We conclude that the likely maximum joint spacing/cartilage thickness of AL 288-1’s hip joint was 2.448 mm which is on par with allometric scaling assumptions (i.e., the smaller bodied AL 288-1 has a more cartilaginous hip joint than the larger bodied human and chimpanzee). Similar estimates were also generated from the single axis translational simulations, despite some implied functional limitations.
The important bit: we cannot ignore translational movement of the hip when estimating the motion-capability of extinct species!
March 2022
HOT OFF THE PRESS! The latest paper just dropped in Scientific Reports. Read the latest blog post here which discusses the paper: https://walkinginthepast.co.uk/the-walking-with-hominins-project-blog/
Whilst this above paper reconstructs musculature in extinct archosaurs, crocodiles and gorillas (a bit different from hominins!), the paper is of great importance for The Walking Hominin project! If we want to build musculoskeletal models of hominins, first we need to reconstuct the musculature and to understand muscle volume and configuration in the body. This new method which we describe here will be used to reconstuct muscles in hominins. How exciting! More to come soon! Sneak peeks & how-to videos likely to drop soon too…
October 2021:
This project began October 2021
Stay tuned for regular updates!
